4.2. Система
капиллярно-кислородного обеспечения сердечной мышцы
4.2.1.
Представление системы
Система
капиллярно-кислородного обеспечения может быть представлена как капиллярно-тканевая
призма, где первичными элементами являются четыре капилляра, приложенное к ним
давление, эритроциты и сердечная ткань. Отношением единства элементов выступает
функциональная зависимость между ними. Законы композиции этой системы будут
представлены нами ниже.
4.2.2.
Особенности капиллярно-тканевого комплекса
Диффузия
кислорода, несмотря на разницу в его потреблении, происходит во всех регионах
сердца в аналогичных условиях. Это связано с постоянством следующих параметров
крови и сердечной ткани млекопитающих.
1. Коэффициент
диффузии кислорода в основном определяется концентрацией воды в органе [284] и
в значительно большей степени его температурой [42]. Концентрация воды в
сердечной ткани постоянна по всему объему сердца [293], а разница температур
для различных слоев сердечной мышцы не превышает 0,5°С [198]. Следовательно, коэффициент диффузии
кислорода практически постоянен по всему объему сердца.
2. Коэффициент
растворимости кислорода в сердечной ткани в умеренной степени зависит от температуры
[209]. Температура во всех регионах сердца различается незначительно,
следовательно, коэффициент растворимости кислорода - постоянная величина по
всему объему сердца.
3. Скорость
диссоциации оксигемоглобина определяется величиной рН и температурой крови. Согласно
анализу экспериментальных данных скорость диссоциации в естественных пределах
изменения величины рН [151, 287] и температуры [175] практически неизменна.
Таким образом, скорость диссоциации оксигемоглобина постоянна по всему объему
сердца.
4. Размеры
эритроцитов различных видов млекопитающих практически одинаковы [174].
5.
Произведение величины площади поверхности эритроцита на их число в единичном
объеме крови млекопитающих - величина постоянная [158].
6.
Концентрация гемоглобина в крови практически одинакова у всех млекопитающих
[164].
7. Величина
среднего напряжения кислорода рО2 во всех регионах сердца одинакова
[213, 216].
Следует
отметить, что перечисленные выше параметры неизменны по всему объему сердца в течение
всего сердечного цикла. Сохраняется также во времени и пространстве постоянство
аспектов архитектоники капиллярного русла в миокарде: неизменность размеров,
параллельность и асимметричность. В то же время абсолютные значения кровотоков
[52, 201] и утилизация кислорода [287] значительно различаются в различных
регионах сердца, например, в верхушке сердца и основании, эндокарде и эпикарде,
левом и правом желудочках. Значительны отличия этих параметров и в отдельные моменты
сердечного цикла. Следует отметить, однако, что одномоментно по объему сердца
большему потреблению кислорода соответствует больший кровоток и большее количество
открытых капилляров. Следовательно, “качественная” картина распределения этих
параметров всюду совпадает. Экспериментально установлено, что для сердца распределение
величин кровотока [261], потребления кислорода [210], расстояний между
открытыми капиллярами [201] и тканевого напряжения кислорода [290] имеет нормальный
характер. Учитывая нормальный закон распределения этих параметров и постоянство
других по объему сердца, можно после усреднения рассматривать капиллярную сеть
как равномерно распределенную по органу с совершенно одинаковыми для
эритроцитов условиями движения и отдачи кислорода.
Усредненное
распределение открытых капилляров в поперечном сечении представляет собою
квадратную решетку (см. рис. 4.1). Функциональной единицей, представляющей
|
|

Рис. 4.1.
Представление открытых капилляров миокарда в виде усредненной квадратной
решетки. А - решетка в золотом режиме гипертензии; Б - решетка в заданном
режиме гипертензии; ![]() (bзс),
(bзс), ![]() (b) - расстояние между капиллярами в золотом и заданном режимах гипертензии.
(b) - расстояние между капиллярами в золотом и заданном режимах гипертензии.
кислородное обеспечение
миокарда, является объем ткани, перфузируемый четырьмя обменными капиллярами.
Эта ячейка имеет форму прямой призмы с квадратным основанием и капиллярами в
качестве боковых ребер (рис. 4.2). Кислородное обеспечение сердечных
|
|

Рис. 4.2. Капиллярно-тканевая призма. а, в - артериальные и венозные концы
капилляров; k - сечение, перпендикулярное капиллярам;
![]() (b) - расстояние между капиллярами (ребрами призмы),
(b) - расстояние между капиллярами (ребрами призмы), ![]() (b)=
(b)= ![]() (bзс)/Öb; lк(b) - длина капилляра, lк(b)=lк(bзс); bзс и b соответствуют золотому и
заданному режимам гипертензии.
(bзс)/Öb; lк(b) - длина капилляра, lк(b)=lк(bзс); bзс и b соответствуют золотому и
заданному режимам гипертензии.
клеток в призме должно
соответствовать следующим двум условиям:
1)кислород
должен поступать в клетки в количестве, адекватном уровню их метаболизма;
назовем этот фактор кислородоснабжением по “количеству”;
2)напряжение
кислорода, поступающего в клетки, должно превышать критическую для данного
уровня метаболизма величину рО2 , ниже которой начинается гибель клеток;
обозначим этот фактор кислородоснабжением по “качеству”.
4.2.3.
Особенности кривой насыщения гемоглобина
По мере
продвижения эритроцита по цепочке сердечных сосудов непрерывно возрастает
разность между значениями HbO2 =97% и рО2 =95 мм рт. ст.
в аорте и значениями pO2 и HbO2 в звеньях цепочки. На
основе кривой насыщения гемоглобина (рис. 3.3) нами была построена разностная
кривая DрО2 =f(DHbO2 ) (рис. 4.3) [152]. При построении этой кривой за
начало отсчета была принята точка с координатами HbO2=97% и рО2=95
мм рт. ст. исходной кривой. В итоге такого построения имеем систему координат,
в которой HbO2 и рО2 поменялись местами, а кривая HbO2=f(рО2)
преобразована “из конца в начало”. Разностная кривая удобна для анализа
закономерностей “разряда” оксигемоглобина на различных участках сосудистой
цепочки. Известно [180], что форма кривых диссоциации оксигемоглобина у разных
видов млекопитающих приблизительно одинакова. Следовательно, подобны друг другу
и разностные кривые. Кривая, представленная на рис. 4.3, построена на основе
кривой диссоциации оксигемоглобина человека, взятой из книги [129]. Кривая
соответствует рН 7,40, pCO2 =39,7 мм рт. ст. и t=37°C; эти параметры крови аналогичны их
естественным величинам в организме.
|
|
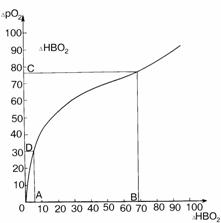
Рис. 4.3. Разностная кривая диссоциации оксигемоглобина человека [148].
По оси абсцисс DHbO2 в %, по оси координат - DpO2 в мм рт. ст.
Интервалы AB и CD соответствуют логариф-мическому участку кривой, где DpO2 =42lgDHbO2 . Интервал AB - DHbO2
=(5,5-68,6)%; интервал CD - DpO2 =30-76 мм рт. ст. Кривая соответствует рН 7,40, pCO2
=39,7 мм рт. ст. и температуре t=37°C .
Произведенный
нами анализ показал, что в пределах DHbO2 =5,5-68,6% и DpO2=30-76 мм рт.
ст. (рис. 4.3, точки A, B, C, D) кривая DpO2=f(DHbО2) представляет собою сегмент логарифмической кривой
DpO2= 42lgDHbO2 .
Всякой
разности двух значений DHbO2 на логарифмическом участке соответствует величина dрО2
dpO2= DрО2(DHbO2*) - DрО2(DHbO2^) = 42lg(DHbO2* /DHbO2 ^),
где DHbO2* , DHbO2^ - любые
значения DHbO2 на логарифмическом участке разностной кривой в пределах
5,5% £ DHbO2 £ 68,6%. Очевидно, что если
величины DHbO2* и DHbO2^ уменьшать в одинаковое число раз, оставаясь при этом в
пределах логарифмического участка, то величина соответствующей им разности dрО2 остается неизменной.
На кривой
диссоциации в ее обычном представлении (см. рис. 3.3) диапазону DpO2 =30-76 мм рт.
ст. соответствует интервал рО2 = 65-19 мм рт. ст., а DHbO2 = 5,5-68,6%
интервал HbO2 = 91,5-28,4%. На артериальном конце капилляра
напряжение кислорода в покое равно 50 мм рт. ст. [61], а на венозном конце - рО2
=20 мм рт. ст. [258]. Таким образом, в золотом режиме гипертензии диапазон рО2(bзс)=50-20 мм рт. ст. полностью
лежит в пределах логарифмического участка кривой диссоциации - 65-19 мм рт. ст.
Напряжение кислорода на венозном конце капилляра совпадает с нижней границей
логарифмического участка, а рО2 на артериальном конце ниже верхней
границы логарифмического участка на 15 мм рт.ст. Рост среднего аортального
давления в b раз вызывает уменьшение разности концентраций кислорода на участках
аорта - артериальный конец капилляра и аорта - венозный конец капилляра в b раз, что равнозначно уменьшению
артерио-венозной разности насыщения на капилляре в b раз [152],
a-vHbO2к (b) = a-vHbO2к(bзс)/b, (4.23)
где a-vHbO2к(bзс), a-vHbO2к(b) - артерио-венозная
разность содержания кислорода в крови капилляра в золотом и заданном режимах
гипертензии. Необходимо отметить, что эти величины постоянны в течение всего
кардиоцикла [256]. Таким образом, при гипертензии в любой момент сердечного
цикла напряжение кислорода на концах капилляра возрастает на одну и ту же
величину dpO2(b)=42lgb,
apO2(b) = 50 + 42lgb, (4.24)
vpO2(b) = 20 + 42lgb, (4.25)
где apO2(b), vpO2(b) - соответственно
напряжение кислорода на артериальном и венозном концах капилляра в заданном
режиме гипертензии. Выражения (4.23)-(4.25) отображают “кислородную” связь
между капиллярами и сердечной тканью. В них представлена картина обеспечения
“качества” кислорода в капилляре. Максимальной гипертензии (b=2) соответствует dpO2=12,6 мм рт.
ст. Отметим, что и при этом смещении величина aрО2(b) не выходит за верхний
предел логарифмического участка кривой насыщения гемоглобина (65 мм рт. ст.).
Таким образом, нижний и верхний пределы изменения рО2 на концах
сердечного капилляра при гипертензии и границы логарифмического участка
практически совпадают друг с другом. Этот феномен, как будет показано ниже,
имеет большое значение для оптимального распределения величин рО2 в
сердечной ткани. Очевидно, что при любой гипертензии артерио-венозная разность
напряжений
a-vpO2(b) = const. (4.26)
Закон (4.26) является
инвариантом системы в покое и при любом уровне гипертензии.
Необходимо
также отметить следующее. Отношение DpO2/DHbO2 плавно
снижается от начала к концу логарифмического, “капиллярного”, участка
разностной кривой (см. рис. 4.3). Эритроцит при движении в любом сечении
капилляра отдает кислород с постоянной скоростью [152]. Если бы эритроцит
перемещался равномерно, то исходя из особенностей разностной кривой величина DpO2/Dlк уменьшалась бы
по направлению от артериального к венозному концу, а DHbO2/Dlк была
постоянной по всей длине капилляра, Однако в действительности эритроцит в
начале капилляра движется быстрее чем в конце [47], поскольку диаметр капилляра
на артериальном конце меньше, чем на венозном на 1-2 мкм [299]. С учетом этой
“коррекции” получается обратная картина - отношение DHbO2/Dlк по направлению
к венозному концу возрастает, а DpO2к/Dlк остается
приблизительно постоянным. Получается, что логарифмический сегмент на капилляре
“выпрямляется”, т.е. имеет место линейная зависимость DpO2к=f(lк).
Этот феномен имеет большое значение для выравнивания “качества” кислородного
обеспечения по всем сечениям капиллярно-тканевой призмы.
Отметим еще
одну особенность кривой насыщения гемоглобина - ее сопряжение с цепочкой сердечных
сосудов, по которой отдельный эритроцит следует от аорты до полости сердца. По
этому признаку кривая насыщения может быть разделена на три сегмента: 1)
артериальный, 2) капиллярный и 3) венозный. Границы этих участков для человека
соответствуют значениям HbO2 , равным 97, 85 и 30%. В соответствии с
таким делением напряжение кислорода по кривой разделяется следующим образом:
артериальный участок - а=95-50=45 мм рт. ст., капиллярный участок - b=50-20=30
мм рт. ст. и “неработающий” участок с=20 мм рт. ст. Для всякой протяженности
длиной d, разделенной на три части - a, b и c, можно вычислить т.н. двойное
отношение,
R=(a+b)(b+c)/cd.
Тройному делению кривой
насыщения человека соответствует R=1,316. В результате анализа кривых насыщения
других видов млекопитающих, представленных в работе [176], по точкам раздела
HbO2 =97%, HbO2 =85% и HbO2 =30% нами получены
величины R=1,23-1,35. Cогласно конформной симметрии отрезки симметричны, если
их двойные отношения равны. С.В.Петухов [103] показал, что если a, b и c
соответственно равны ряду чисел 1,618, 1,6182 и 1,6183,
то R=2,618/2=1,309; эта величина обозначена С.В.Петуховым золотым вурфом.
Очевидно, что величины R=1,23-1,35 весьма близки к золотому вурфу.
Следовательно, кривые насыщения различных видов млекопитающих, имея инвариантом
золотой вурф, симметричны друг другу.
4.2.4.
Капиллярно-тканевая призма в режимах гипертензии
Представим
взаимное расположение мышечного волокна и питающих его капилляров в виде прямой
квадратной призмы А с четырьмя перфузируемыми капиллярами в качестве ребер (см.
рис. 4.4). форма призм одинакова как в систолу, так и в диастолу при любой
гипертензии. Начнем анализ кислородного обеспечения призм с золотого режима
гипертензии. Длина ребер-капилляров постоянна в течение всего кардиоцикла.
Разница между призмами состоит лишь в том , что расстояние между капиллярами в
“диастолической” призме в ![]() раз больше, чем в
“систолической”; Bs(bзс), Bd(bзс) -
раз больше, чем в
“систолической”; Bs(bзс), Bd(bзс) -
|
|
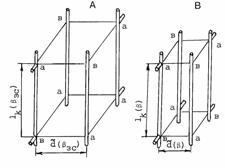
Рис. 4.4. Капиллярно-тканевая призма в режимах гипертензии. А - призма
в золотом, Б - призма в заданном режиме гипертензии. lк(b) - длина капилляра, lк(b)=lк(bзс); ![]() (b),
(b), ![]() (bзс) - расстояние
между капиллярами в золотом и заданном режимах гипертензии,
(bзс) - расстояние
между капиллярами в золотом и заданном режимах гипертензии, ![]() (b)=
(b)= ![]() (bзс)/Öb.
(bзс)/Öb.
соответственно среднее
потребление кислорода единичным объемом сердечной мышцы за систолу и диастолу в
золотом режиме гипертензии. Легко показать, что произведение объема
систолической призмы на скорость потребления кислорода единичным объемом призмы
равно аналогичному “диастолическому” произведению. Очевидно, что потребление
кислорода объемами систолической и диастолической призм за единицу времени будет
одинаковым. Необходимо при этом учесть, что величины рО2 на концах
капилляров [256] и величина рО2 в отдельной сердечной клетке [289]
постоянны в течение кардиоцикла. Следовательно, распределение рО2 в
сечениях призм, равноудаленных от основания, будет одинаковым и в систолу, и в
диастолу. При этом нет оснований считать, что существуют различия в
распределении рО2 в различных регионах сердца. Установлено [187,
213], что удельное кровоснабжение 100 г ткани в правом желудочке за единицу
времени составляет 60% от такового в левом желудочке. Аналогичное соотношение
для желудочков имеет место и по удельной скорости потребления кислорода [287].
Показано, что среднее внутриклеточное рО2 в обоих желудочках
одинаково и равняется 9,6 мм рт. ст. [213, 216]. Очевидна аналогия
распределения рО2 в призмах по всему сердцу. Необходимо отметить при
этом, что и удельное кровоснабжение, и удельное потребление кислорода, и
плотность капиллярной сети желудочков соотносятся между собою по золотому
сечению!
Рассмотрим
преобразование призмы, соответствующей одной из фаз (например, систоле), в
режимах гипертензии (см. рис. 4.4). При заданном уровне гипертензии b призма А превращается в
призму Б. В этой призме расстояние между ребрами уменьшается в Öb раз [149], а потребление
кислорода единичным объемом призмы за единицу времени увеличивается в b раз [146]. Очевидно, что
потребление кислорода объемами систолических призм А и Б за единицу времени
одинаково; та же величина потребления кислорода имеет место и для
диастолических призм А и Б. Очевидно, что скорость потребления кислорода
призмами постоянна в любой момент времени при всех уровнях гипертензии. Иначе
говоря, с “кислородной” точки зрения все призмы в течение сердечного цикла и
при всяком уровне гипертензии эквивалентны друг другу, т.е. потребляют одно и
то же количество кислорода. Если бы при гипертензии величины рО2 на
артериальном и венозном концах капилляров сохранялись неизменными, то
распределение величин рО2 в сечениях призм, равноудаленных от
основания, оставалось бы постоянным при любом уровне гипертензии. В
действительности же показано, что при росте гипертензии величина рО2
в миокарде возрастает [114, 257], что и должно быть в соответствии с выражениями
(4.24) и (4.25).
Велико
значение оптимального сопряжения асимметричности капилляров и логарифмичности
“капиллярного” участка кривой насыщения для обеспечения “качества” кислорода по
всему объему призмы. Равномерность обеспечения “качества” по объему призмы
связана со следующими обстоятельствами. Четыре капилляра по ребрам призмы
расположены таким образом, что 1) кровотоки в противоположных капиллярах имеют
обратные направления; 2) концы каждого капилляра по отношению к двум соседним
смещены на полдлины капилляра вверх и вниз. За счет замедленного движения
эритроцита в капилляре логарифмический участок кривой насыщения преобразуется
по отношению к длине капилляра в прямую. За счет этого феномена обеспечивается
линейное распределение рО2 по длине капилляра (рис. 4.5). В золотом
режиме благодаря условиям 1) и 2), а также линейному распределению pO2
по длине капилляров, в каждом сечении призмы сумма рО2 в капиллярах
приблизительно равна постоянной величине
SpO2к(bзс) = 140 мм рт. ст. (4.27)
|
|
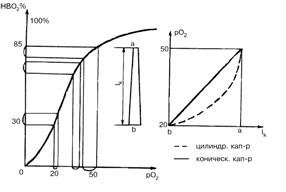
Рис. 4.5. “Распрямле-ние” разностной кри-вой DpO2=f(DHbO2) на
капилляре. lк- длина капилляра, а, в - артериальный и венозный концы
капилляра.
При переходе
от одного сечения к другому приращения величины рО2 в противоположных
капиллярах одинаковы, но обратны по знаку. Вследствие этого обстоятельства и
того, что в каждом сечении призмы потребление кислорода одинаково,
распределение рО2 во всех сечениях практически аналогично.
Произведем анализ распределения рО2 в некотором k-сечении призм А и
Б, равноудаленном от основания. Распределение рО2 в k-сечении в
золотом режиме соответствует кривой 1 (рис. 4.6). В некотором режиме гипертензии
напряжение кислорода в капиллярах для k-сечения призмы Б увеличивается на
величину dpO2 =42lgb; вследствие этого кривая
плотности распределения рО2 призмы Б смещается на ту же величину
(кривая 2, рис. 4.6).
|
|
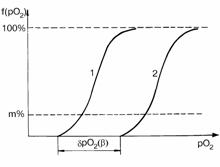
Рис. 4.6. Плотность распределе-ния величин pO2 в k-сечении
капиллярно-тканевой призмы. По оси абсцисс - pO2 (условные
едини-цы); по оси ординат - долевое распределение величин pO2 в
k-сечении призмы (в %). 1 - распределение pO2 в золотом, 2 -
распределение pO2 в заданном режиме гипертензии; dpO2 =42lgb - приращение pO2
на концах капилляров при переходе от покоя к заданному режиму гипертензии.
Естественно,
что подобное же смещение имеет место во всех сечениях призмы Б. Тем самым
обеспечивается “качество” кислорода, адекватное для заданного уровня гипертензии
и постоянное по всему объему призмы. Скорость отдачи кислорода эритроцитом в каждом
сечении призмы линейно зависит от разности рО2 в эритроците и потребляющей
среде [230]. Поскольку в каждом сечении приращения dpO2(b) одинаковы и в капиллярах,
и в ткани, то как следствие этого, постоянна и скорость отдачи кислорода
движущимся эритроцитом. Эта скорость неизменна по всей длине сердечного капилляра
в любой момент кардиоцикла и при любой гипертензии. Она имеет, как мы уже знаем,
максимально значение
U(b) = 215%/c.
Эта величина
является инвариантом всего микроциркуляторного русла сердца. Смысл его заключается
в том, что любое сечение всякого микрососуда в любой момент времени независимо
от уровня гипертензии отдает кислород с постоянной, максимально возможной
скоростью.
Покажем
симметрийность распределения величин рО2 в мышечном волокне. Рассмотрим
с этой целью некоторую последовательность тканевых призм вдоль мышечного
волокна (рис. 4.7). Нельзя не обратить внимания на то, что сечения, в которых
“координатное”, точечное распределение рО2 совершенно аналогично,
повторяются через каждые 2lк ; в этом случае имеет место симметрия
параллельного сдвига. С другой стороны, координатное распределение рО2
в любом сечении через каждые 1/2lк “поворачивается” на 90°, т.е. имеет место поворотная симметрия.
Таким образом, распределение рО2 вдоль волокна происходит в
соответствии с винтовой симметрией; каждой величине рО2 соответствует
“своя” винтовая линия с шагом h=2lк . Благодаря этому обстоятельству
ни одна из клеток по длине мышечного волокна не находится в менее благоприятных
условиях, чем остальные. Более того, все клетки находятся в одинаковых
“кислородных” условиях. Шаг h=2lк является инвариантом продольного
распределения рО2 по волокну в любой момент сердечного цикла и при
всякой гипертензии.
Важно при этом
отметить и симметрийность распределения рО2 в “диффузионной” цепочке
капилляров. Нетрудно убедиться в том, что здесь имеет место зеркальная симметрия
и симметрия параллельного сдвига. Относительно любого соединения концов
капилляров (артериального или венозного) существует зеркальная симметрия, когда
по обе стороны соединения значения рО2 зеркально повторяются. С
другой стороны, через каждые (2n+1)lк, где n=1, 2, ... и т.д.,
происходит параллельный сдвиг значений рО2 по цепочке капилляров.
т.е. величины рО2 в капиллярах повторяются. Кроме того, для четырех
цепочек капилляров, параллельных волокну, также существует свой вид симметрии.
Каждая из них может быть совмещена с любой из оставшихся за счет двух параллельных
сдвигов - в горизонтальном и вертикальном направлениях. Наличие различных видов
симметрии в отдельной и совокупности цепочек позволило обеспечить винтовое
распределение рО2 вдоль мышечного волокна, что равносильно
постоянству “качества” кислородного обеспечения для всех мышечных клеток.
Таким образом,
на примере тканевой призмы с ребрами-капиллярами нами проанализированы оба
механизма кислородного обеспечения клеток - по “количеству” и “качеству”. Эти
механизмы взаимосвязаны друг с другом. Первый механизм в условиях гипертензии
обеспечивается увеличением плотности капиллярной сети в b раз, а второй -
возрастанием давления на капилляре также в b раз. Как следствие этого, кровоток в
капиллярной сети увеличивается в b2 раз. Оба механизма
рефлекторно связаны с увеличением среднего аортального давления в b раз. Наличие этих
гемодинамических факторов, а также логарифмичность участка на разностной
кривой, взаимная асимметрия и параллельность капилляров, позволяет обеспечить
адекватное снабжение миокарда кислородом при любом уровне гипертензии.
Произведем
расчет “цены” транспорта единичного объема кислорода через капиллярный участок
в режимах гипертензии.
Кровоток и
давление на уровне капиллярного русла равны постоянным величинам в любой момент
сердечного цикла. При гипертензии по отношению к золотому режиму кровоток через
капиллярный участок возрастает в b2 раз, а давление на
капиллярах - в b раз [147, 149, 152]. Средний расход мощности на перфузию капилляров за
сердечный цикл в некотором режиме гипертензии
Wпк(b) = (1/T(bзс)/b2)![]() b2qк(bзс,t)dt = b3Wпк(bзс),
b2qк(bзс,t)dt = b3Wпк(bзс),
где Wпк(bзс), Wпк(b) - средняя за кардиоцикл
перфузионная мощность в генерации капилляров в золотом и заданном режимах
гипертензии, pк(bзс,t), qк(bзс,t) - мгновенные значения
давления и кровотока в капиллярах в золотом режиме гипертензии.
Поскольку
объем капиллярного русла при гипертензии возрастает в b раз, а
bк(b) = b2bк(bзс),
то
Wхк(b) = b3Wхк(bзс),
где Wхк(bзс), Wхк(b), bк(bзс), bк(b) - соответственно средняя
за сердечный цикл суммарная химическая мощность в генерации капилляров,
скорость расхода химической энергии в единичном объеме капилляра в золотом и
заданном режимах гипертензии.
Естественно,
что
SWк(b) = b3SWк(bзс), (4.28)
где SWк(bзс), SWк(b) - общий расход
механической и химической энергии в капиллярном русле за единицу времени в
золотом и заданном режимах гипертензии.
Поскольку
общий расход мощности в капиллярном русле увеличивается в b3 раз, а скорость потребления
кислорода сердечной мышцей - в b2 раз [146], то очевидно, что с ростом гипертензии
“цена” доставки диффундируемого в капилляре кислорода возрастает в b раз. Самый “дешевый”
транспорт кислорода в капиллярах имеет место в золотом режиме гипертензии (b=bзс=1), а наибольшая “цена” доставки
соответствует режиму максимальной гипертензии (b=2). Значит, и для
артериального, и для капиллярного участков коронарного русла наиболее и менее
всего благоприятные режимы транспорта кислорода совпадают. Если рассматривать
генерации микрососудов, из которых кислород диффундирует в ткани, то окажется,
что самый “дешевый” кислород поступает из капилляров в золотом режиме
гипертензии (покое). На капиллярном участке в сердечную мышцу поступает 85% от
всего кислорода, утилизированного миокардом. Если учесть, что покой составляет
наибольшую часть протяженности жизни животного, то очевидно, что кислородное
снабжение сердца млекопитающих обеспечивается в основном при минимальной
“цене”.
В заключение
следует отметить инварианты, присущие капиллярно-тканевым призмам в течение
сердечного цикла при каждом уровне гипертензии:
1)неизменность
пространственной ориентации капилляров (параллельность и асимметрия);
2)постоянство
скорости потребления кислорода объемом призмы в целом;
3)постоянство
“винтового” распределения рО2 вдоль мышечного волокна;
4)постоянство
разности рО2 на артериальном и венозном концах капилляров;
5)постоянство
скорости отдачи кислорода.
Наряду с инвариантами,
представленными в (4.20) и (4.21) для “гемодинамической” цепочки, эти инварианты
составляют оптимальную основу организации “диффузионной” цепочки капилляров.
Совокупность установленных нами инвариантов обусловливает оптимальность
крово-кислородного обеспечения миокарда на капиллярном уровне.